英茂盛业生物科技有限公司
023-67630383
023-67630383
 销售咨询QQ2548969917 技术咨询QQ1291769782
销售咨询QQ2548969917 技术咨询QQ1291769782热线/微信:13681365274
销售咨询QQ2548969917 技术咨询QQ1291769782微信扫一扫
下载说明书


目标蛋白在细菌宿主中形成包涵体对获得大量生物活性蛋白质造成很大困难。从包涵体中获得生物活性蛋白需要大量手工工作,并且重组蛋白的产量通常很低。这篇文章中,作者回顾了该领域的发展,旨在通过优化过程的各个步骤,特别是包涵体的溶解和溶解蛋白的重折叠,提高重组蛋白的产量和质量。
温和溶解方法基于对包涵体聚集体中的蛋白质分子具有天然样结构这一事实的理解。这种方法在溶解包涵体的同时,保留天然样蛋白质结构。随后通过蛋白质再折叠和纯化获得高回收率的生物活性蛋白质。文章还讨论了影响从包涵体中回收生物活性蛋白的其它参数,并且提出了用于生物活性蛋白质的高通量回收的温和增溶方法的效用的示意性模型。
重组蛋白在大肠杆菌中的高水平表达通常导致表达的蛋白分子聚集成包涵体。在蛋白质表达过程中使用高温、高诱导物浓度和在强启动子系统下表达通常导致所需蛋白质以高翻译速率表达。这耗尽了细菌蛋白质质量控制系统,部分折叠和错误折叠的蛋白质分子聚集形成包涵体。细菌胞质溶胶的还原环境、缺乏真核分子伴侣和翻译后机制也有助于包涵体的形成。虽然近年来已报道细菌包涵体提供了许多生物技术应用,但本综述的重点是阐述从包涵体中回收重组蛋白,特别是使用温和的溶解过程。
包涵体的形成给使用大肠杆菌作为宿主的重组蛋白的生产和纯化带来了很大的障碍。包涵体需要大量加工,包括从细胞中分离、溶解、重折叠和纯化以产生生物活性蛋白质。尽管在理解包涵体中蛋白质的结构细节方面有了新的发展,但大多数时候,溶解和重折叠是以经验方式进行的,导致功能性蛋白质的回收率很低。包涵体蛋白质的溶解和它们的再折叠可以根据关于包涵体蛋白质的结构和功能特征的新信息进行微调。在大肠杆菌中,由于蛋白质聚集的特异性,包涵体主要由目标重组蛋白组成。如果能够开发出从包涵体聚集体中回收正确折叠的蛋白质分子的方便有效的方法,将减少对大量色谱纯化步骤的需要。包涵体具有天然构象的蛋白质分子,一些包涵体具有显著的生物活性。使用高浓度的离液剂如尿素和盐酸胍(GdnHCl)会导致这些现有的二级结构完全变性,并经常导致蛋白质分子在重折叠过程中聚集。
据报道,与用高浓度离液剂溶解相比,使用温和的溶解策略从包涵体中回收和再折叠蛋白质效率更高。这些温和的增溶剂在一定程度上保留了蛋白质现有的二级结构;在重折叠过程中抑制蛋白质聚集,从而提高生物活性蛋白质的回收率。本综述的目的是更新关于包涵体中蛋白质结构特征的最新发现,以及从大肠杆菌包涵体中高通量回收生物活性蛋白质的温和增溶方法的新进展。
影响大肠杆菌包涵体形成的因素
多种因素有助于蛋白质聚集物作为包涵体的形成。在大肠杆菌中,包涵体在细胞内积累为折射颗粒,典型尺寸范围为0.2至1.5微米。在电子显微镜下,它们看起来是具有光滑或粗糙表面形态的致密、可折射颗粒。一般来说,包涵体是球形的,但也可以是圆柱形或卵形,以适应细菌细胞。在大肠杆菌中,它们大多位于细菌细胞的一极或两极。最近有报道称,细菌细胞两极包涵体的定位是由细菌胞质溶胶中的大分子拥挤驱动的。在细胞分裂期间,只有一个子细胞接受包涵体,而在另一个细胞中包涵体的形成从头开始。一般来说,包涵体聚集在胞质溶胶中,但据报道,具有分泌信号的蛋白质也在周质空间形成聚集体。
重组蛋白的表达涉及多种合成途径,并在转录和翻译水平受到调控。通常,当蛋白质的表达水平超过总细胞蛋白质的2%时,会导致包涵体的形成。有利于重组蛋白高速表达的因子增加了表达蛋白聚集成包涵体的机会。高拷贝数的目的基因、强启动子系统和高诱导物浓度有利于包涵体的形成。拷贝数过高会使细胞的代谢负担增加,而拷贝数过低会带来质粒在后代中丢失的风险。载体的启动子强度在调节蛋白质的表达水平方面也起着重要作用。当具有强启动子系统的细菌细胞被诱导产生重组蛋白时,它们经历了代谢负担水平的提高,表达的重组蛋白聚集成包涵体的机会增加。细菌胞质溶胶的还原环境也会导致蛋白质聚集。包涵体的形成也依赖于蛋白质的氨基酸序列,高度疏水的蛋白质有更多的机会聚集成包涵体。包涵体中存在细菌伴侣蛋白已被广泛报道。已经表明,缺少胞质伴侣,ClpB增加易聚集重组蛋白的不溶性表达。蛋白质聚集在大多数类型的细胞中是一种受调节的现象。在大肠杆菌中,这些包涵体形式的聚集体仅沉积在两极。这种极性定位导致子细胞之间包涵体以不对称的方式分布。这有助于提高没有聚集物的子细胞的细胞生长率,并使细菌更适于进一步分裂。
包涵体中蛋白质的结构特征
King及其同事首次对包涵体的形成进行了详细的机理研究。他们将噬菌体P22尾丝蛋白的体外折叠与体内折叠途径进行比较,获得了一个蛋白折叠模型。该模型描述了多肽从部分折叠的中间体到折叠和聚集途径的分配。人们普遍认为,蛋白质聚集是一个高度特异性的过程,蛋白质分子只能与相同或高度相关的其他蛋白质分子聚集。包涵体聚集体在本质上也是高度特异性的,因为重组蛋白构成了这些聚集体的主要部分。Kopito及其合作者在2001年使用两种不同的具有聚集倾向的蛋白,用不同的荧光蛋白标签标记,使用荧光共振能量转移(FRET)显示这些蛋白在真核细胞中不共聚集。通过用两种不同的荧光标签[蓝色荧光蛋白(BFP)和绿色荧光蛋白(GFP)]表达淀粉样β蛋白,然后进行FRET分析,进一步证实了细菌包涵体中蛋白质聚集的特异性。据观察,相同类型的蛋白质分子在细胞中共聚集,而不同的蛋白质如VP1不与淀粉样β蛋白共聚集。这些结果表明,包涵体中的蛋白质聚集是一种高度特异的现象。
包涵体在本质上是高度动态的,参与包涵体形成的蛋白质分子可以可逆地解聚并折叠成其天然形式。聚集在包涵体中的蛋白质分子与其可溶性对应物之间存在动态平衡。因此,包涵体的连续构建和解构过程发生了。其中聚集和可溶形式的蛋白质分子可以自由交换它们的定位。
传统上,包涵体被认为是无定形类型的聚集体,缺乏任何结构规则性。在过去的十年中,许多报道提供了支持包涵体淀粉样性质的证据,尽管最近表明包涵体可能在性质上不是淀粉样。包涵体聚集体已被证明与淀粉样蛋白特异性染料结合,如硫黄素T和刚果红。傅立叶变换红外(FTIR)光谱分析显示,包涵体富含β含量,并给出与交叉β结构相对应的信号,这是淀粉样蛋白聚集的特征。这些聚集体中交叉β结构的存在也从包涵体给出的X射线衍射图中明显可见,该包涵体与淀粉样纤维产生的包涵体具有惊人的相似性。淀粉样蛋白聚集是一种基于成核的现象,蛋白质的聚集物可以作为相似蛋白质聚集的种子。这也适用于大肠杆菌包涵体。据报道,像淀粉样聚集体一样,包涵体可以在体外引发类似蛋白质的聚集。与可溶性蛋白质一起温育的包涵体已被证明与长纤维结构相关,该长纤维结构与淀粉样纤维具有形态相似性。还报道了使用H/D交换NMR发现包涵体聚集体中淀粉样蛋白的形成是一种残基特异性现象。即使是一个点突变也能抑制包涵体的形成过程。有几个在线服务器预测具有形成淀粉样蛋白高倾向的区域。Hamodrakas及其同事最近应用这种工具来预测负责包涵体形成的多肽区域。
据报道,细菌包涵体具有天然的二级结构。白细胞介素-1β、β-内酰胺酶、人粒细胞集落刺激因子(GCSF)等蛋白质形成具有天然二级结构的包涵体。报告天然二级结构存在的初步研究主要使用FTIR光谱。固态核磁共振波谱也被用于证明包涵体中存在天然结构。此外,大量包涵体已被证明具有相当大的生物活性。虽然1989年就有第一份报告表明包涵体中存在活性,但直到最近十年,活性包涵体才因其在各种过程中的潜在应用而受到关注。β-内酰胺酶、β-半乳糖苷酶和GCSF的包涵体已被证明具有相当大的生物活性。具有生物活性的包涵体聚集体被称为非经典包涵体。大多数非经典包涵体的特点是蛋白质分子排列松散,因此即使在低浓度的变性剂下也能溶解。包涵体中活性分子的比例取决于蛋白质表达过程中的物理条件。包涵体的质量可以通过改变表达温度来调节。虽然也有在高温下形成活性包涵体的报道,但一般认为低表达温度有利于非经典包涵体的形成。
活性包涵体的形成可以通过用合适的接头将蛋白质连接到易于聚集的肽上来诱导。Wu等人证明了将自组装离子肽ELK16 (LELELKLK)2连接到蛋白质可以导致形成具有显著生物活性的包涵体。类似肽的表面活性剂也被用于形成生物活性包涵体。还报道了全长绿色荧光蛋白作为聚集诱导物的附着,以形成活性包涵体,以及连接肽的重要性。
包涵体聚集体具有高度动态的性质。除了重组蛋白,包涵体还包含细菌膜、其他宿主蛋白和RNA的成分。如前所述,包涵体含有淀粉样原纤维或原纤维。Riek和他的同事使用核磁共振光谱表明,包涵体中的一部分蛋白质的H/D交换率不同于淀粉样蛋白。这表明包涵体中存在结构异质性。这种异质性可能是由于污染的宿主蛋白。还有一种可能性是,未折叠、部分折叠或甚至天然结构的重组蛋白分子可以共存于包涵体中,并部分导致包涵体中蛋白聚集体的结构异质性。这也为一些包涵体中活性的存在提供了解释。目前包涵体的理论模型假设它们主要由淀粉样蛋白样原聚集体组成。其他构象(包括天然状态)的重组蛋白分子被捕获在淀粉样原纤维或原纤维的网络中[11]。重组蛋白的淀粉样成分和其他结构形式的比例取决于许多因素,包括表达过程中使用的物理条件[53]。所有这些新信息强烈表明,包涵体蛋白具有相当数量的天然样结构。
从包涵体中回收生物活性蛋白质
由于蛋白质分子在包涵体中处于聚集状态,溶解包涵体并将溶解的蛋白质重折叠成生物活性形式是一项具有挑战性的任务。从包涵体中纯化蛋白质的常规策略由四个主要步骤组成:纯化包涵体的分离、包涵体的溶解、溶解的蛋白质的重折叠和通过各种色谱技术纯化重折叠的蛋白质。包涵体的溶解和溶解蛋白分子的复性是从包涵体中回收生物活性蛋白的最关键步骤。
从大肠杆菌细胞中分离纯化的包涵体
包涵体是高度特异性的聚集体,主要由感兴趣的重组蛋白组成。因此,在溶解和重折叠之前,有必要将包涵体聚集体分离和纯化至均一。从包涵体中纯化蛋白质减少了对标签和多个色谱步骤的需要。如上所述,表达条件影响包涵体的质量。低表达温度有助于形成柔软、非经典的包涵体,这些包涵体可以用非变性溶剂溶解。据报道,在表达过程中调节pH会影响包涵体的质量。用于从细菌细胞中分离包涵体的方法包括使用超声或French press的机械细胞破裂法和使用细胞裂解剂如溶菌酶的化学细胞破裂法。据报道,细胞破碎方法的选择对包涵体的质量有很大影响。与机械方法如超声或匀浆相比,通过化学方法进行细胞破碎被认为是更好的。后者影响包涵体的质量,并导致最初属于可溶性部分的蛋白质分子聚集。还报道了使用机械和化学细胞破碎技术的合适组合。由于包涵体与其他细胞成分相比密度较高,因此可以通过离心从全细胞裂解物中分离出来。错流膜微滤也用于从宿主细胞蛋白质中分离包涵体]。分离的包涵体含有几种杂质,如宿主蛋白、RNA和宿主膜碎片。使用几个洗涤步骤进一步纯化它们。用低浓度的去污剂如脱氧胆酸和Triton X-100 洗涤不仅有助于获得高度纯化的包涵体,还有助于去除膜碎片。据报道,非经典包涵体对纯化时的pH非常敏感,因为在纯化过程中使用高pH会导致蛋白质分子溶解。蔗糖密度梯度超速离心也被用于包涵体的纯化。纯包涵体有助于减少重折叠过程中污染物的干扰,并减少对大量纯化步骤的需求。
包涵体蛋白质的溶解
传统上,使用高浓度的变性剂和离液剂如尿素和盐酸胍(GdnHCl)溶解包涵体。对于含有多个半胱氨酸残基的蛋白质,在这些增溶剂中加入β-巯基乙醇或二硫苏糖醇,以减少不正确的二硫键。使用高浓度离液剂溶解包涵体导致蛋白质结构的完全破坏。在某些情况下,这导致蛋白质分子在重折叠过程中聚集。由于包涵体聚集体已被证明具有类似天然的二级结构并可能具有活性,因此使用“温和”的溶解过程是有利的,该过程不会完全展开这些类似天然的蛋白质结构。
如前所述,包涵体本质上是动态的,在折叠和聚集的蛋白质分子之间存在平衡。这一事实可用于在非变性缓冲液中溶解包涵体,无需任何增溶剂的帮助。N-乙酰基-d-氨基葡萄糖2-差向异构酶的包涵体已经用Tris–HCl缓冲液在pH 7下溶解。据报道,溶解的蛋白质具有活性。使用温和的溶解条件可以从非经典包涵体中提取生物活性蛋白质分子。该过程保留了包涵体中存在的天然样蛋白质结构,因此绕过了重折叠步骤。低浓度的有机溶剂(如5%正丙醇和DMSO)和去污剂(如0.2% N-月桂酰肌氨酸)已被用作非经典包涵体溶解的温和增溶剂。在许多情况下,低浓度尿素也被用于溶解包涵体聚集体。这些增溶剂导致活性重组蛋白的提取,而不需要任何重折叠步骤。这些方法的主要缺点是它们局限于经典的包涵体。
为了有效溶解包涵体,已经开发了几种温和的溶解方法,在溶解过程中保持蛋白质分子处于部分折叠状态。使用这些方法,溶解的蛋白质分子的重折叠从部分折叠的形式开始,抑制分子在重折叠过程中进入聚集途径。图1给出了这些温和增溶方法的示意图。使用碱性pH、高压、去污剂、有机溶剂和低浓度离液剂的温和溶解过程已经用于从包涵体中回收生物活性蛋白质。表1总结了这些方法。在大多数情况下,包涵体分离的改进和现代重折叠方法与温和溶解相结合的使用提高了生物活性蛋白质的总回收率。
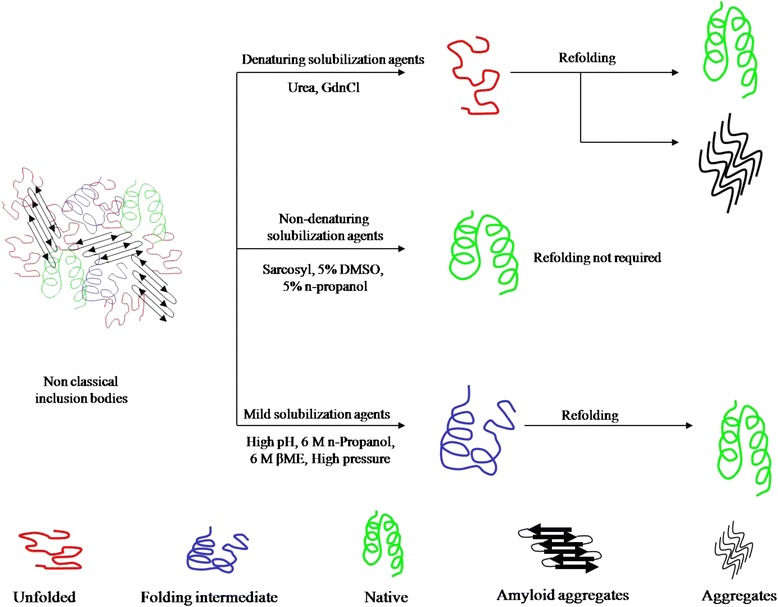

高静水压也有助于包涵体聚集体的溶解[64]。高压(2-4千巴)破坏了分子间的相互作用并分解了包涵体。去除施加的高压导致蛋白质分子的重新折叠[64,67]。在9°C下施加2.4千巴的压力溶解包涵体,随后在0.4千巴和20°C下重折叠,也显示出改善重组内皮抑制素的重折叠[68]。
据报道,使用极端pH值的缓冲液也是一种温和的溶解方法。高pH (>12)缓冲液结合2 M尿素已成功用于溶解包涵体[19,61]。已经描述了高pH缓冲液通过将天然样二级结构保持在溶解状态来提高重折叠产量[19]。低浓度尿素的增溶作用[63],去污剂如N-月桂酰抗坏血酸和月桂酰-L-谷氨酸盐也被报道可以提高包涵体中生物活性蛋白的产量[65,66,69]。
据报道,有机溶剂如β-巯基乙醇(βME) [70]和正丙醇与低浓度尿素结合使用是一种提高蛋白质回收率的新型增溶策略[60]。这两种溶剂都被用于溶解人生长激素包涵体。在人生长激素的情况下,使用基于正丙醇的缓冲液通过保留天然二级结构而提高了重折叠产量,超过了常规使用的基于尿素或GdnHCl的缓冲液[60]。有机溶剂,主要是醇类,长期以来被认为与蛋白质相互作用并影响其二级和三级结构。许多醇也显示出对蛋白质二级结构具有稳定作用,并且已知甚至在非结构化肽中诱导螺旋性[71,72]。因此,使用有机溶剂溶解包涵体聚集体为基于尿素/GdnHCl的溶解提供了可行的替代方案。据报道,多种蛋白质被正丙醇和2 M尿素的混合物溶解[60]。这为使用基于有机溶剂的溶解来提高从细菌包涵体中回收蛋白质开辟了新的可能性。
没有一种单一的增溶方法适用于每一种蛋白质,因此,必须筛选增溶剂以优化增溶。为了快速方便地筛选增溶剂,Hahn及其同事报道了一种基于浊度的高通量分析方法,用于筛选大量增溶剂[73]。因此,通过理解引起蛋白质聚集的主导力来适度溶解包涵体聚集物是成功开发大肠杆菌包涵体高通量蛋白质重折叠方法的关键。
溶解蛋白的再折叠
溶解的包涵体蛋白质通过除去增溶剂而重新折叠。在重折叠缓冲液中稀释溶解的蛋白质[59]和在重折叠缓冲液存在下透析溶解的蛋白质[74]是用于回收功能活性蛋白质的最常用方法。这些方法的缺点是需要高缓冲液,特别是对于大规模操作,以及由于蛋白质聚集导致的低重折叠产率。在重折叠过程中,低蛋白浓度和减少的分子间相互作用是抑制蛋白聚集和增加重折叠产率的先决条件。为了实现这一点,开发了一种称为脉冲稀释的改良稀释方法,这种方法显著降低了缓冲液需求,并提高了蛋白质的重折叠产量[54]。
为了提高重折叠蛋白质的质量和扩大工业应用的规模,已经开发了填充色谱柱中的重折叠。不同的色谱方法已被用于溶解蛋白质的再折叠。尺寸排阻[75-79]、离子交换[80-83]和亲和层析[84-86]是最广泛使用的方法,而疏水相互作用层析[87,88]也已用于某些情况。这种被称为柱复性的方法与传统的稀释和透析方法相比有几个优点。使用大小排阻色谱进行重折叠导致在用重折叠缓冲液洗脱的过程中,折叠形式与聚集和错误折叠形式分离。在其他色谱方法中,重折叠过程发生在蛋白质固定在固体支持物上之后,导致重折叠单元的空间分离和分子间相互作用的降低。因此,色谱床中的重折叠过程可以在高蛋白质浓度下进行。此外,重折叠过程伴随着变性剂的去除和目标蛋白的纯化。这减少了纯化过程中涉及的步骤数量,并使该过程自动化[89,90]。有报道称使用固定在层析介质上的伴侣蛋白来模拟体内折叠[91-95]。尽管观察到重折叠产率增加,但由于高加工成本,工业应用受到限制。该领域的持续发展已经改进了该方法,并且该方法的最新进展旨在增加重折叠产率[96]和优化重折叠条件以提高蛋白质的质量[97]。
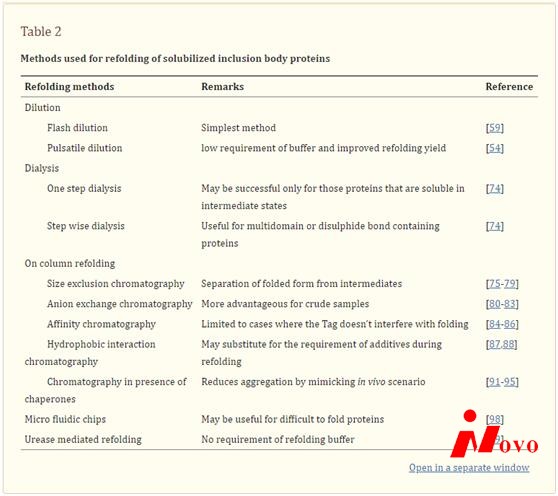
最近描述了一种新的重折叠方法,包括使用微流控芯片[98]。在微流控芯片中,变性剂的浓度通过控制微通道中层流的扩散来维持,并且已经用于难以折叠的蛋白质如柠檬酸合酶的重折叠。还描述了另一种使用脲酶进行蛋白质重折叠的方法,该方法包括通过脲酶催化反应从溶解的蛋白质溶液中逐渐去除尿素[99]。这种系统的一个主要优点是可以在低体积下实现有效的蛋白质重折叠,而无需使用重折叠缓冲液,这可以显著降低生产成本。表2总结了用于可溶性包涵体蛋白重折叠的不同方法。
用于重折叠的方法的成功部分取决于重折叠缓冲液的组成。重折叠过程中最常见的问题是目标蛋白的聚集,大多数重折叠策略包括在重折叠缓冲液中使用某些添加剂来抑制聚集。除了聚集抑制剂,某些折叠增强剂也用于提高产量。重折叠缓冲液中常用的添加剂包括离液剂,如低浓度的尿素[19]或盐酸胍[74],氨基酸,如甘氨酸[100],精氨酸[101,102]和脯氨酸[103,104],多元醇和糖,如聚乙二醇[105-108],甘油[104,109],山梨醇[110]和蔗糖[104]。非去污剂两性离子,如磺基甜菜碱、取代的吡啶和吡咯以及酸取代的氨基环己烷也在重折叠过程中用作添加剂[111-114]。这些添加剂总结在表3中。

已知一些蛋白质在体内的重折叠需要在信号肽和多肽成熟部分之间存在N-末端肽原[115]。一些体外重折叠策略利用在重折叠缓冲液中添加前肽来增加重折叠产率[116-118]。类似地,受体内蛋白质折叠启发的其他策略利用伴侣蛋白对溶解的蛋白质进行再折叠。伴侣是天然的蛋白质折叠辅助机械。它们通过依次捕获和释放重折叠中间体并阻止它们相互作用来发挥作用。使用伴侣蛋白和其他折叠催化剂,如肽基脯氨酰基顺反异构酶或蛋白质-二硫键异构酶,加速折叠途径的限速步骤,已显示可提高重折叠产率[91-95]。洗涤剂胶束作为人工伴侣系统已被用于提高复性过程的产量。在这里,蛋白质折叠中间体与去污剂相互作用,形成混合的蛋白质-去污剂胶束,并防止分子间相互作用。中间形式的再折叠是通过添加环糊精来启动的,环糊精通过形成更稳定的去污剂环糊精复合物而从蛋白质-去污剂复合物中剥离去污剂[119,120]。最近,基于使用与智能聚合物结合的亲和配体分离蛋白质,描述了纯化和重折叠蛋白质的策略[121-124]。这些聚合物在提供特定刺激如温度、pH或离子强度变化时沉淀,从而沉淀出感兴趣的蛋白质。然后通过逆转沉淀条件从聚合物中回收结合的蛋白质。没有用于最佳蛋白质重折叠的通用缓冲液,必须针对每种情况单独筛选重折叠缓冲液的组成[9,125-126]。
结论
先进的结构技术大大增强了我们对包涵体聚集体中蛋白质结构的理解。包涵体现在被认为具有构象异质性,淀粉样结构构建了一个网络,其中具有其他构象的蛋白质分子,包括天然构象的蛋白质分子被捕获。已经开发了温和的增溶方法,以保存这些天然样结构,并为常规使用高浓度离液剂提供可行的替代方案。折叠蛋白质的有效回收也减少了大量色谱步骤的负担。结合改进的重折叠方法,温和的溶解导致从包涵体中高通量回收生物活性蛋白。从包涵体中回收蛋白质的过程包括四个步骤:
1 .包涵体纯化至均质(小心裂解细胞,通过去污剂洗涤/超速离心纯化包涵体)
2 .使用温和增溶剂(碱性pH/流体静压/有机溶剂基缓冲液/2-3 M尿素/去污剂)溶解包涵体
3 .用最佳重折叠缓冲液
4.重折叠蛋白的纯化。这些步骤可以被优化以从大肠杆菌的包涵体中回收大量的生物活性蛋白质。
Microb Cell Fact. 2015; 14: 41.
Published online 2015 Mar 25. doi: 10.1186/s12934-015-0222-8

